In this revision, James D. Paruk, David C. Evers, and Jay Mager revised all sections of the account. Peter Pyle contributed to the Appearance page. Michael A. Patten contributed to the Systematics page.
Gavia immer
(Brünnich, 1764)
PROTONYM:Colymbus Immer
Brünnich, 1764. Ornithologia borealis, sistens collectionem avium ex omnibus, Imperio Danico subjectis (etc.), p.38.
UPPERCASE: current genusUppercase first letter: generic synonym● and ● See: generic homonymslowercase: species and subspecies●: early names, variants, misspellings‡: extinct†: type speciesGr.: ancient GreekL.: Latin<: derived fromsyn: synonym of/: separates historical and modern geographic namesex: based onTL: type localityOD: original diagnosis (genus) or original description (species)
Dedicated in honor of Jud Dayton— conservationist, proud Minnesotan, and longtime member of the Cornell Lab of Ornithology's Administrative Board.
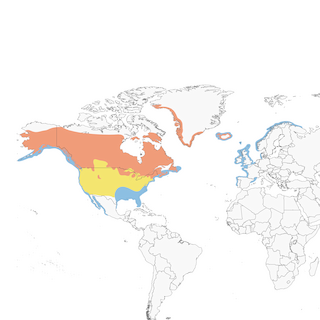
One of five loon species worldwide, the Common Loon is an iconic symbol of lake country across the Nearctic, well known for its distinctive and haunting vocalizations. It is a predominately piscivorous diving bird that nests on oligotropic lakes, but also occupies mesotrophic lakes at the southern periphery of its range. Breeding lakes vary widely in size, from 5 hectares (supporting 1 pair) to 92,000 hectares (supporting over 100 pairs). It breeds across Canada and Alaska (typically not in tundra), south to the northern border of the contiguous United States, and in coastal Greenland and Iceland. During the nonbreeding period, it predominately occurs along coasts, but also frequently on freshwater reservoirs. In North America, it mainly winters from Alaska to the Baja Peninsula and Gulf of California, and from Newfoundland to Florida, and the northern Gulf of Mexico. In Europe, it winters in the North Sea, off coasts of Norway, Sweden and the United Kingdom, and the Celtic Sea, from the Republic of Ireland to southern France and Spain.
The Common Loon is a long-lived species (rarely, > 35 years) with delayed maturity and low fecundity. In parts of its range, behavior (1
Piper, W. H., K. B. Tischler, and M. Klich (2000). Territory acquisition in loons: the importance of take-over. Animal Behaviour 59: 385–394.
, 2
Paruk, J. D. (2006). Testing hypotheses of social gatherings of Common Loons (Gavia immer). Hydrobiologia 567: 237–245.
, 3
Mager, J. N., C. Walcott, and D. Evers (2007). Macrogeographic variation in the body size and territorial vocalizations of male Common Loons (Gavia immer). Waterbirds 30(1): 64–72.
, 4
Mager, J. N., C. Walcott, and W. H. Piper (2007). Male Common Loons, Gavia immer, communicate body mass and condition through dominant frequencies of territorial yodels. Animal Behaviour 73: 683–690.
, 5
Mager, J. N., C. Walcott, and W. H. Piper (2012). Male Common Loons signal greater aggressive motivation by lengthening terrtiorial yodels. Wilson Journal of Ornithology 124: 74–81.
, 6
Mager, J. N., and C. Walcott (2014). Dynamics of an aggressive vocalization in the Common Loon (Gavia immer): a review. Waterbirds 37 (Special Publication 1): 37–46.
, 7
Piper, W. H., J. N. Mager, C. Walcott, L. Furey, N. Banfield, A. Reinke, F. Spilker, and J. A. Flory (2015). Territory settlement in Common Loons: no footholds but age and assessment are important. Animal Behaviour 104: 155–163.
, 8
Piper, W. H., K. M. Brunk, G. L. Jukkala, E. A. Andrews, S. R. Yund, and N. G. Gould (2018). Aging male loons make a terminal investment in territory defense. Behavioral Ecology and Sociobiology 72: 95.
), physiology (9
Haefele, H. J., I. Sidor, D. C. Evers, D. E. Hoyt, and M. A. Pokras (2005) Hematologic and physiologic reference ranges for free-ranging adult and young common loons (Gavia immer). Journal of Zoo and Wildlife Medicine 36: 385–390
), population dynamics (10
Mitro, M. G., D. C. Evers, M. W. Meyer, and W. H. Piper (2008). Common Loon survival rates and mercury in New England and Wisconsin. Journal of Wildlife Management 72: 665–673.
, 11
Paruk, J. D., M. D. Chickering, D. Long, H. Uher-Koch, A. East, D. Poleschook, V. Gumm, W. Hanson, E. M. Adams, K. A. Kovach, and D. C. Evers (2015). Winter site fidelity and winter movements in Common Loons (Gavia immer) across North America. Condor 117(4): 485–493.
, 12
Piper, W. H., K. M. Brunk, J. A. Flory, and M. W. Meyer (2017). The long shadow of senescence: age impacts survival and territory defense in loons. Journal of Avian Biology 48: 1062–1070.
), movements (13
Piper, W. H., J. D. Paruk, D. C. Evers, M. W. Meyer, K. B. Tischler, M. Klich, and J. J. Hartigan (1997). Local movements of color-marked Common Loons. Journal of Wildlife Management 61(4): 1253–1261.
, 14
Kenow, K. P., M. W. Meyer, D. C. Evers, D. C. Douglas, and J. Hines (2002). Use of satellite telemetry to identify Common Loon migration routes, staging areas and wintering range. Waterbirds 25(4): 449–458.
, 15
Kenow, K. P., D. J. Hoffman, R. K. Hines, M. W. Meyer, J. W. Bickham, C. W. Matson, K. R. Stebbins, P. Montagna, and A. Elfessi (2008). Effects of methylmercury exposure on glutathione metabolism, oxidative stress, and chromosomal damage in captive-reared Common Loon (Gavia immer) chicks. Environmental Pollution 156(3): 732–738.
, 16
Gray, C., J. D. Paruk, C. R. DeSorbo, L. J. Savoy, D. E. Yates, M. Chickering, R. B. Gray, K. M. Taylor, D. Long IV, N. Schoch, W. Hanson, and D. C. Evers (2014). Strong link between body mass and migration distance for Common Loons (Gavia immer). Waterbirds 37: 64–75.
, 17
Paruk, J. D., D. Long IV, S. L. Ford, and D. C. Evers (2014). Common Loons wintering off Louisiana coast tracked to Saskatchewan during the breeding season. Waterbirds 37: 47–52.
), and threats (18
Paruk, J. D., J. N. Mager, and D. C. Evers (2014c). Introduction: An Overview of Loon Research and Conservation in North America. Waterbirds 37: 1–5.
) have been well characterized through long-term research programs that often include capture and marking of individuals. Recent advances in high-resolution population models (10
Mitro, M. G., D. C. Evers, M. W. Meyer, and W. H. Piper (2008). Common Loon survival rates and mercury in New England and Wisconsin. Journal of Wildlife Management 72: 665–673.
), habitat quality ranking models (19
Evers, D. C., M. Sperduto, C. Gray, J. D. Paruk, and K. Taylor (2019). Restoration of Common loons following the North Cape Oil Spill, Rhode Island, USA. Science of the Environment 695: 133849.
), and mercury wildlife criterion value models (20
Schoch, N., A. K. Jackson. M. Duron, D. C. Evers, M. J. Glennon, C. T. Driscoll, X. Yu, H. Simonin, and A. K. Sauer (2014). Wildlife Criterion Value for the Common Loon (Gavia immer) in the Adirondack Park, New York, USA. Waterbirds 37: 64–75.
) have provided refined quantitative tools to support science-based management and policy decisions. These models have contributed to a better understanding of the primary threats to the species. Further efforts to mark and track individuals (e.g., satellite telemetry), sample for contaminant and genetic profiles, and translocate chicks (21
Kneeland, M., V. Spagnuolo, M. Pokras, J. Paruk, N. Schoch, D. C. Evers, V. Stout, L. Attix, K. Silber, and A. Dalton (2020). A novel method for captive rearing and translocation of juvenile common loons. Zoo Biology 39(4): 263–270.
) will enhance long-term, landscape-scale conservation.
Overall, populations are relatively healthy and robust, with an estimated global breeding population of 260,000 territorial pairs. Inclusion of the nonbreeding cohort increases the estimated total adult population to 624,000 individuals, a number that increases to 780,000 during fall migration, with inclusion of juveniles. Approximately 30 percent of the fall population migrates to the Pacific coast of North America and 70 percent to the Gulf of Mexico and Atlantic coasts. About 94 percent of the global population resides in Canada, where the species is relatively protected from shoreline development and recreational activities, though other threats could impact Canada’s robust populations.
Widely recognized as an indicator of aquatic health, populations may be adversely impacted by anthropogenic threats in both breeding and wintering ranges. Concerns during the breeding period include mercury pollution (22
Evers, D. C., J. D. Kaplan, M. W. Meyer, P. S. Reaman, W. E. Braselton, A. Major, N. Burgess, and A. M. Scheuhammer (1998). Geographic trend in mercury measured in Common Loon feathers and blood. Environmental Toxicology and Chemistry 17(2): 173–183.
, 23
Meyer, M. W., D. C. Evers, J. J. Hartigan, and P. S. Rasmussen (1998). Patterns of Common Loon (Gavia immer) mercury exposure, reproduction, and survival in Wisconsin, USA. Environmental Toxicology and Chemistry 17(2): 184–190.
, 24
Kenow, K. P., S. Gutreuter, R. K. Hines, M. W. Meyer, F. Fournier, and W. H. Karasov (2003). Effects of methyl mercury exposure on the growth of juvenile common loons. Ecotoxicology 12(1–4): 171–182.
, 25
Burgess, N. M., and M. W. Meyer (2008). Methylmercury exposure associated with reduced productivity in Common Loons. Ecotoxicology 17(2): 83–91.
, 26
Evers, D. C., L. J. Savoy, C. R. DeSorbo, D. E. Yates, W. Hanson, K. M. Taylor, L. S. Siegel, J. H. Cooley, M. S. Bank, A. Major, K. Munney, B. F. Mower, H. S. Vogel, N. Schoch, M. Pokras, M. W. Goodale, and J. Fair (2008). Adverse effects from environmental mercury loads on breeding Common Loons. Ecotoxicology 17(2): 69–81.
, 27
Evers, D. C., K. A. Williams, M. W. Meyer, A. M. Scheuhammer, N. Schoch, A. T. Gilbert, L. Siegel, R. J. Taylor, R. Poppenga, and C. R. Perkins (2011). Spatial gradients of methylmercury for breeding common loons in the Laurentian Great Lakes region. Ecotoxicology 20: 1609–1625.
, 28
Tozer, D., C. Falconer, and D. Badzinski (2013). Common loon reproductive success in Canada: the west is best but not for long. Avian Conservation and Ecology 8(1).
), and in the southern part of the range, recreational activities and human disturbance (29
Windels, S. K., E. A. Beever, J. D. Paruk, A. R. Brinkman, J. E. Fox, C. C. MacNulty, D. C. Evers, L. S. Siegel, and D. C. Osborne (2013). Effects of water-level management on nesting success of Common Loons. Journal of Wildlife Managment 77: 1626–1638.
, 30
Grade, T. J., M. A. Pokras, E. M. LaFlamme, and H. S. Vogel (2018). Population-level effects of lead fishing tackle on Common Loons. Journal of Wildlife Management 82: 155–164.
). During migration, botulism outbreaks (31
Kenow, K. P., S. C. Houdek, L. J. Fara, B. R. Gray, B. R. Lubinski, D. J. Heard, M. W. Meyer, T. J. Fox, and R. J. Kratt (2018). Distribution and foraging patterns of Common Loons on Lake Michigan with implications for exposure to type E avian botulism. Journal of Great Lakes Research 44: 497–513.
) are a major concern, and oil spills (32
Sperduto, M. B., S. P. Powers, and M. Donlan (2003). Scaling restoration to achieve quantitative enhancement of loon, seaduck, and other seabird populations. Marine Ecology Progress Series 264: 221–232.
, 33
Paruk, J. D., D. Long IV, C. Perkins, A. East, B. J. Sigel, and D. C. Evers (2014). Polycyclic Aromatic Hydrocarbons detected in Common Loons wintering off coastal Louisiana. Waterbirds 37: 85–93.
, 34
Paruk, J. D., E. M. Adams, H. Uher-Koch, K. A. Kovach, D. Long IV, C. Perkins, N. Schoch, and D. C. Evers (2016). Polycyclic aromatic hydrocarbons in blood related to lower body mass in common loons. Science of the Total Environment 565: 360–368.
, 19
Evers, D. C., M. Sperduto, C. Gray, J. D. Paruk, and K. Taylor (2019). Restoration of Common loons following the North Cape Oil Spill, Rhode Island, USA. Science of the Environment 695: 133849.
), marine wind farms (35
Mendel, B., P. Schwemmer, V. Peschko, S. Muller, H. Schwemmer, M. Mercker, and S. Garthe (2019). Operational offshore wind farms and asociated ship traffic cause profound changes in distribution patterns of Loons (Gavia spp.). Journal of Environmental Management 231: 429–438.
), and emaciation syndrome (36
Forrester, D. J., W. R. Davidson, R. E. Lange, R. K. Stroud, L. L. Alexander, J. C. Franson, S. D. Haseltine, R. C. Littell, and S. A. Nesbitt (1997). Winter mortality of Common Loons in Florida coastal waters. Journal of Wildlife Diseases 33(4): 833–847.
) can be problematic in the wintering period. How populations will respond to climate change remains largely unknown (37
Bianchini, K., D. C. Tozer, R. Alvo, S. P. Bhavsr, and M. L. Mallory (2020). Drivers of declines in common loon (Gavia immer) productivity in Ontario, Canada. Science of the Total Environment 738: 139724
, 38
Piper, W. H., J. Grear, B. Hoover, E. Lomery, and L. M. Grenzer (2020). Plunging floater survival causes cryptic population decline in the Common Loon. Condor: Ornithological Applications 122: 1–10.
). The health and sustainability of Common Loon populations depends on our response to minimize known threats across their range.
Paruk, J. D., D. C. Evers, J. W. McIntyre, J. F. Barr, J. Mager, and W. H. Piper (2021). Common Loon (Gavia immer), version 2.0. In Birds of the World (P. G. Rodewald and B. K. Keeney, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.comloo.02
Partnerships
A global alliance of nature organizations working to document the natural history of all bird species at an unprecedented scale.